2015/11/19
【概要】
植物は、微生物の侵入や感染を監視するため、微生物に特有の構成成分(細菌のべん毛タンパク質フラジェリンやカビの細胞壁構成成分キチンなど)と特異的に結合する免疫センサー※1を細胞表面に配置しています。そのような微生物センサー※2が標的成分を感知すると、補助タンパク質(BAK1)※3とともに免疫センサー複合体を形成して微生物の感染を抑える免疫応答を誘導します。しかし、微生物センサーは植物の成長を助ける良性の共生菌や常在菌がもつ標的成分も認識することから、排除すべき悪性の病原菌を識別するために植物は別のセンサーも活用していると予想されますが、その詳細は不明でした。
奈良先端科学技術大学院大学(学長:小笠原直毅)バイオサイエンス研究科植物免疫学研究室の西條雄介准教授らは、共同研究者である山田晃嗣博士(現京都大学)らとともに、悪玉の微生物による細胞のダメージを感知、識別して植物免疫が強化される仕組みを明らかにしました。人為的に微生物センサーの補助因子BAK1を除去して微生物センサーによる免疫応答を低下させた植物では、そのような細胞の異常を感知する細胞ダメージセンサー※4の働きが活発になっていたのです。これまで西條准教授らはモデル植物のシロイヌナズナを材料に、植物自身のダメージを感知するセンサーが植物の生体防御に重要であることを世界に先駆けて明らかにしており、その裏付けとなる研究成果です。
また、西條雄介准教授らは研究の過程で、世界中で甚大な作物被害を起こしている「炭疽病菌」がBAK1を除去して感染を進めること、また感染の拡大阻止に植物の細胞ダメージセンサーが重要な役割を担っていることを突き止めました。
同様の仕組みはヒトを始めとした動物にもあると推測されます。例えば腸内細菌のような常在菌とヒトの細胞にダメージを引き起こす病原菌を識別する際に働いていると予想されます。さらに、病原菌が侵入して生存の危機が迫った際に初めて植物が免疫力を高める仕組みの発見は、拮抗しがちな植物成長と病害抵抗性を両備する農作物の開発にも弾みをつけるものです。
本成果は、平成27年11月16日(月)付で欧州科学誌The EMBO Journalにオンライン掲載されました。
【説明】
植物も動物と同様に優れた免疫システムを備えており、病原菌の侵入・感染をいち早く察知して的確に防御応答を展開することでその脅威から身を守っています。これまでに、①植物細胞の表面には微生物センサーが複数種類あり、それらが微生物の構成因子(細菌のべん毛タンパク質やカビの細胞壁構成成分など)を認識すると防御応答が誘導されること、②病原菌は微生物センサーの働きを妨害することで感染を成立させることが知られていました。しかしながら、動物のように異物に特異的な抗体を産生できない植物が多様な病原菌を無害な常在菌と識別する仕組みや、病原菌による妨害にもかかわらず免疫応答を活性化してその感染の拡大を防ぐ仕組みは不明でした。
西條准教授の研究グループは、以前の研究で、細胞ダメージ警報として働くと考えられる免疫活性化因子PROPEPペプチドとそのセンサーPEPR※5が、微生物センサーによる免疫応答の増幅や全身への伝播に果たす重要性を明らかにしてきました。しかしながら、微生物センサーとPEPRの情報伝達系が連携する仕組みや、PROPEPペプチド(またはそのPepと呼ばれる、免疫活性化能をもつ部分配列)がダメージ警報として働く仕組みは不明のままでした。今回、共同研究グループの山田晃嗣博士(現京都大学)らを中心としてこれら未解決の問題を解き進め、植物が病原菌を識別する仕組みで鍵となる部分を解明することに成功しました。
【実験の手法】
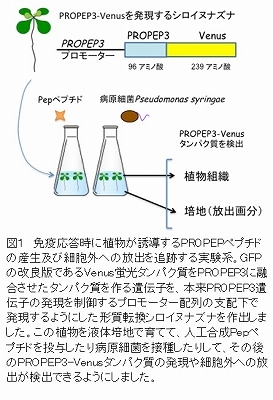
PEPRも細菌フラジェリンのセンサーFLS2も、それぞれ特異的な標的因子を認識すると、直ちにBAK1を含むセンサー複合体を形成して情報伝達を行います。シロイヌナズナの変異体を用いて、BAK1遺伝子の欠損によるPEPR情報伝達系への影響や、BAK1とPEPRを同時に欠損させた変異体植物において様々な免疫応答への影響を解析しました。また、蛍光タンパク質GFPの改良版であるVenusを融合させたPROPEP3ペプチドがPROPEP3遺伝子の誘導時に発現するようにした形質転換植物を作製して、病原菌の接種後PROPEPペプチドの産生や放出が追跡できるようにしました(図1)。
【実験の結果】
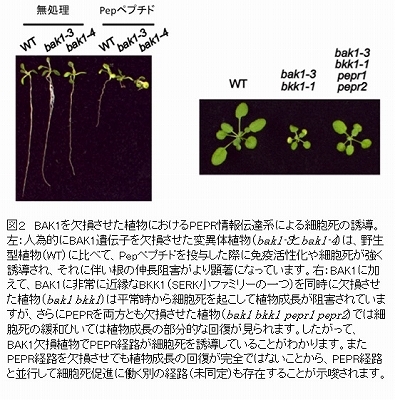
BAK1遺伝子を欠損した植物にPepペプチドを合成して投与すると、PEPRによる情報伝達系が細胞死を増強する方向に働くことが判明しました(図2)。これまでにBAK1が細胞死を抑制する働きも持ち、その抑制効果を除去した植物では細胞死が進みやすいことが知られています。今回の結果は、PEPR情報伝達系がBAK1無しでも機能すること、並びにBAK1という細胞死ブレーキが働かない環境でオンになると細胞死を亢進することを示すものです。次にPep投与により発現が変動する遺伝子を網羅的にプロファイリングした結果、植物の免疫ホルモンであるサリチル酸の応答がBAK1欠損植物で強化されていることがわかりました。また、BAK1欠損植物は、FLS2による免疫応答が低下しているにもかかわらず、野生型植物と比べて同等もしくはそれ以上の抵抗性を複数の病原菌に対して示します。しかし、BAK1に加えて2種類あるPEPRを同時に欠損させた植物では抵抗性が著しく損なわれることが確かめられました。すなわち、FLS2などの微生物センサーの働きが低下したBAK1欠損植物において、PEPR経路がバックアップとして働くことで植物免疫が維持・強化されていることが明らかになりました。さらにPROPEP3-Venusの追跡実験により、病原菌の病原性や感染時に起こる細胞死の度合いに応じてPROPEPペプチドの産生や細胞外への放出が増強され、BAK1欠損植物では放出が一層促進されることがわかりました。したがって、PROPEPペプチドが病原菌感染に伴う細胞ダメージの警報として働き、病原菌の識別に寄与することが確実となりました。
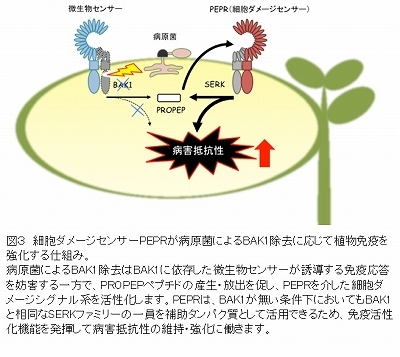
さらに、炭疽病を引き起こすColletotrichum属糸状菌(カビ)が植物に感染すると、BAK1タンパク質を選択的に除去して増殖を図ることが示されました。これに対して、植物はBAK1除去時に活発に働くPEPR情報伝達系を動員することで炭疽病菌の感染拡大を阻止することが明らかとなりました(図3)。
【本研究の意義】
植物の免疫応答は病原菌の撃退に寄与する一方で植物の成長には阻害的に働くなど、過度に活性化されると植物の生存を不利にする面もあるため、病原菌に対して適切な規模で発動されるよう緻密に制御されています。しかし、その仕組みは未だによくわかっていません。植物細胞の表面には、動物と同様に、ロイシンリッチリピート(LRR)と呼ばれるタンパク質構造によって細胞外の標的因子を検出するセンサーが多種類あり、植物の発生や環境応答に重要な役割を果たしています。FLS2やPEPRのような免疫センサーに加えて、細胞間でやり取りする植物ホルモンや情報ペプチドを認識するセンサーがあり、その多くが補助タンパク質BAK1を必要とします。そのため、BAK1は病原菌が植物の細胞プロセスを妨害して感染しようとする際に格好の狙い目となってしまいます。今回の研究により、病原菌によるBAK1除去という感染戦略に対して、免疫応答を強化したり細胞死を進めて警報を全身に増幅・拡散したりする仕組みが明らかになりました。このPEPR情報伝達系は高等植物に広く共有されており、今回の研究でキュウリやイチゴ等の農作物に甚大な被害を起こす炭疽病菌に対して特に有効であることも示されました。元来あらゆる病原菌の感染は細胞ダメージにつながり、それに応じて免疫を活性化する経路は炭疽病菌のみならず広範な病原菌に対して必要な際に必要な強度の免疫応答を発動するのに重要です。さらに研究が進むことで、植物の成長を維持しながら人為的に植物免疫をコントロールする技術の開発につながることも期待されます。
【用語解説】
※1 免疫センサー:体内に侵入した異物を認識すると免疫応答を誘導するタンパク質の一群で免疫受容体とも呼ばれています。それぞれの免疫センサーが特異的に認識する異物(標的因子)と免疫センサーはちょうど鍵と鍵穴のような関係にあります。植物は、細胞表面で微生物に特有の構成成分をそれぞれ特異的に認識する免疫センサーを持っています。また、病原菌が植物細胞内に注入して感染を促進する因子や感染に伴う細胞異常を感知する免疫センサーを細胞内にも持つことで病原菌の認識に役立てています。植物は、動物のように遺伝子組み換えを起こして多様な抗体タンパク質を生み出せません。その代わりに、細胞表面センサーで微生物の侵入を感知し、さらに病原菌が引き起こす細胞異常を細胞内センサーで監視する二段構えのシステムによって、一定数の免疫センサーで環境中の多様な微生物に対応していると考えられています。
※2 微生物センサー:免疫センサーのうち、良性や悪性かを問わず微生物に特有の構成成分を認識するもので、植物には、細菌のべん毛タンパク質フラジェリンを認識するFLS2やタンパク質合成装置の一部を認識するEFR、また糸状菌(カビ)の細胞壁の構成成分であるキチンオリゴ糖を認識するCERK1などが知られています。これまで同定された植物の微生物センサーは全て細胞膜結合型のタンパク質であり、細胞表面において細胞外スペース中の標的成分を検出すると考えられています。一方、動物も同様の微生物センサーを細胞内にも持つことが知られています。
※3 補助タンパク質BAK1:細胞外領域にロイシンリッチリピート(LRR)と呼ばれるタンパク質構造を持ち、細胞内領域にリン酸化酵素(プロテインキナーゼ)を持つ、LRR受容体キナーゼ(LRR-RK)と総称される膜貫通型のタンパク質の一つ。植物は同様な構造をもつLRR-RKを多種類持ち(シロイヌナズナの約27000強の遺伝子のうち200以上がLRR-RKを作る遺伝子)、その多くが細胞外の標的因子(微生物の構成成分や植物細胞間でやり取りする情報因子等)を感知するセンサーとして働くと考えられています。これらの細胞外センサーは標的因子が結合すると細胞内に情報を伝達して細胞プロセスを調節するが、その際に補助因子としてBAK1を必要とするものが多く知られています。シロイヌナズナにはBAK1に非常によく似たタンパク質が他に4種類あり、LRR-RKの大ファミリーの中でSERKと総称される小ファミリーを形成していて、互いの機能の一部を相補し合える関係にあります。今回の研究によって、PEPR情報伝達系の補助因子としてのBAK1の役割は他のSERKメンバーが十分に補える一方で、細胞死のブレーキとしてのBAK1の役割は他のSERKメンバーが十分に補えないことが分かりました。
※4 細胞ダメージセンサー:細胞の傷害ダメージや細胞異常(通常行われるはずのプロセスが行われないなどの異変)を感知するセンサータンパク質の総称。植物において、細胞傷害の感知に役立つと考えられているセンサーには、細胞壁の破砕成分を認識するセンサーや今回の研究対象であるPEPR等が知られています。上述の、病原菌の感染に伴う細胞異常を検出する細胞内の免疫センサーも機能的にはまさに細胞ダメージセンサーの一種と見なせます。今回の研究では、細胞表面で働く微生物センサーと細胞ダメージセンサーがBAK1除去をきっかけに織りなす連携の仕組みについて解明を進めました。
※5 PROPEPペプチドとそのセンサーPEPR:PROPEPペプチドはおよそ100アミノ酸から成るペプチドで、C末端に免疫活性化能をもつPepと呼ばれるおよそ20アミノ酸の配列を持っています。Pep配列をもつペプチドはシロイヌナズナには8種類存在し、イネやダイズなどの高等植物にも広く存在することが知られています。PROPEPペプチドは、細胞外にタンパク質を分泌する通常の経路に運ばれるためのタグ配列を持たないため、平常時には細胞内にとどまり、細胞膜にダメージが生じた際に細胞外に放出されると考えられています。人工合成したPep配列を植物に投与すると免疫応答が誘導され、その際にPep配列を認識するセンサーとして働くのがPEPRと呼ばれるLRR-RKであり、シロイヌナズナには2種類存在します。今回の研究によって、病原菌感染時にPROPEPペプチドおよびその派生ペプチドが産生・放出されること、さらにPEPRによって認識されるために必要な細胞外への放出がBAK1欠損条件で増大することが明らかになりました。
【関連リンク】
・タイトル
Danger peptide receptor signaling in plants ensures basal immunity upon pathogen‐induced depletion of BAK1
・論文は以下に掲載されております。
DOI:http://dx.doi.org/10.15252/embj.201591807
naistar:http://hdl.handle.net/10061/10462
(NAIST Academic Repository:naistar)
・以下は論文の書誌情報です。
Kohji Yamada, Misuzu Yamashita‐Yamada, Taishi Hirase, Tadashi Fujiwara, Kenichi Tsuda, Kei Hiruma, Yusuke Saijo; The EMBO journal, 201591807, published online 16 November 2015
【本研究内容についてコメント出来る方】
川崎 努 教授
近畿大学農学部バイオサイエンス学科 植物分子遺伝学研究室
TEL: 0742-43-7335 FAX: 0742-43-7335 E-mail: t-kawasaki@nara.kindai.ac.jp
高野 義孝 准教授
京都大学大学院農学研究科 応用生物科学専攻 植物病理学分野
TEL: 075-753-6133 FAX: 075-753-6131 E-mail: ytakano@kais.kyoto-u.ac.jp
【本プレスリリースに関するお問い合わせ先】
奈良先端科学技術大学院大学 バイオサイエンス研究科 植物免疫学研究室 西條 雄介
TEL:0743-72-5690 FAX:0743-72-5559 E-mail:saijo@bs.naist.jp